
Do Prokaryotes Have Circular DNA?
Recent research has revealed that some prokaryotes may have as many as four linear or circular chromosomes, in contrast to the majority of prokaryotes, such as E. coli, which have a single circular DNA molecule that makes up their whole genome. For instance, the cholera-causing bacteria Vibrio cholerae has two circular chromosomes.

What is circular DNA, and does it exist in prokaryotes? We know that eukaryotic cells have multiple linear chromosomes, but do prokaryotes have circular DNA? In this article, we will learn about circular DNA and plasmids. We will also discuss rotaxanes and catenanes. This article will answer both questions and more.
Eukaryotic cells have multiple linear chromosomes.
Chromosomes are DNA molecules that contain tens of thousands of base pairs. Eukaryotic cells contain large, linear chromosomes, while prokaryotic cells contain smaller circular ones. Some cells contain multiple types of chromosomes, including chloroplasts and mitochondria. Here are some common types of chromosomes found in cells. If you’re unsure which type your cell has, look at these examples.
The eukaryotic cell has many pairs of linear chromosomes, and each pair has its unique form. In interphase, eukaryotic chromosomes become less compact, allowing transcription to occur. This looser structure is known as chromatin, and it has important implications for the expression of genes. Chromosomes are composed of hundreds of thousands of base pairs and are about 10,000 times shorter than taut without proteins.

Bacteria and eukaryotic cells have multiple linear lines, and their circular chromosomes are scattered throughout their taxonomic groups. Interestingly, linear chromosomes are thought to have evolved due to large-scale lateral gene transfer. In the case of bacteria, mitochondrial genes were transferred to the developing nuclear genome and spliced into the circular chromosome, which created multiple copies of the host chromosome. Parasite jumping genes also played a crucial role in this evolution.
Rotaxanes
The molecular architecture of rotaxanes is composed of a dumbbell-shaped molecule threaded through a ring-like macrocycle. Rotaxane is derived from the Latin for “wheel” and “axle.” Rotaxanes are trapped molecules because their ends are more significant than the internal diameter of the ring. Thus, unthreading the rotaxane would require considerable distortion of covalent bonds.
AHL pore opens at +140 mV, and rotaxane formation begins. During co-translocation, the rotaxane remains associated with the pore. After the rotaxane is formed, it undergoes retro-translocation. The co-translocational unfolding phase is recorded as the potential increases from negative to positive.
Rotaxanes in prokaryote microbial cells have several biomedical uses. These dyes can be used in optical bioimaging and can be modified to act as cellular transport agents and drug delivery systems. The chemical reactions are best carried out in weakly polar organic solvents and highly polar protic solvents. They are highly stable and a potential biomedical tool shortly.
Catenanes
The massively catenated structure of DNA is vital for maintaining genetic information and allowing genetic exchange, an essential part of sexual reproduction. To link catenanes, a chain of DNA is broken. It then passes through the closed circle and re-closes, forming a catenane. This complex process is carried out by enzymes known as topoisomerases.
Replication initiates at a single replication origin and terminates at a site opposite the origin. The replication terminator region consists of two polar domains, Ter and Tus, and this complex counteracts the activity of the helicases to terminate the process. As a result, DNA replication forks pause in predictable patterns during the process, allowing the ssDNA to pass through the break.
Viruses and plasmids also employ this process to copy nucleic acids. Again, molecular markers such as EM and electrophoresis determine how the substrates are converted. Once the nicks are repaired, the molecule produces the second daughter strand. The process is then repeated for a second time, with the un-nicked strand serving as a template.
Telomeres
Telomeres protect the chromosome ends of bacteria. These protective structures allow the chromosomes to replicate without losing their genetic information. Most prokaryotes have circular chromosomes instead of telomeres, which allows bacteria to preserve their genetic information. However, oxidative stress is a significant threat to telomeres.
Cells divide 50 and 70 times before death, and telomeres get shorter with each division. This reduction in length is referred to as the Hayflick limit. If a cell’s telomeres shorten, it inhibits cell division and produces senescence. To learn more about telomeres, visit the Telomerase Database and Wikimedia Commons.
The length of telomeres varies between different prokaryotes, from micro-zoa to eukaryotes. In eukaryotes, telomere length depends on the age of the organism. In prokaryotes, telomere length is measured in kilobases and micro-zoa. Telomere length differs from one organism to another, which means that the size of the chromosome largely determines telomere length.
Cellulose
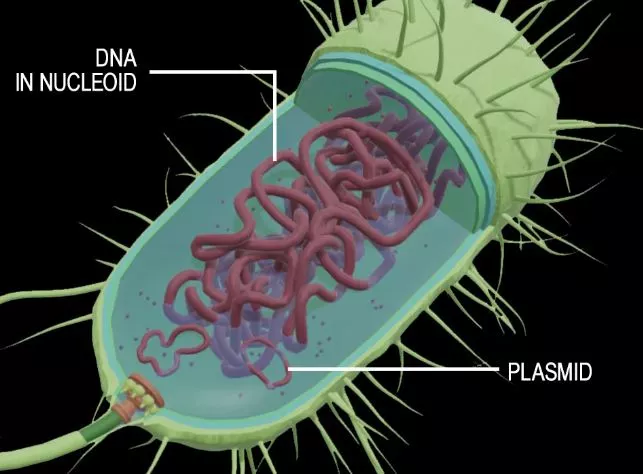
The cellular structure is composed of cellulose, and circular DNA is called a prokaryotic cell. This cell form contains the genetic material and the cell chemicals but lacks a defined nucleus and organelles. These cells are made up mostly of bacteria and are characterized by small free-floating ribosomes. Circular DNA is found inside each prokaryotic cell. The circular DNA is paired with plasmids. In addition, prokaryotes do not have a cell wall made of cellulose or chitin, but their cell walls are rigid.
In addition to their cellulose and circular DNA, prokaryotic cells also contain small replicons, which are normal constituents of prokaryotes. They can multiply in different types of cells without needing a large replicon. They are the primary participants of horizontal gene exchange and are underestimated by most biologists. They play a significant role in the reproduction of prokaryotic cells.
These prokaryotic cells have numerous advantages over eukaryotic cells. Their small size permits the production of enormous numbers of different cells, which increase the chances of cross-feeding and cell exchange. These two essential aspects of life in prokaryotes are critical in maintaining the stability of the biosphere. All animals’ soil and alimentary tracts harbor a diverse community of prokaryotic cells.
Eukaryotic cells have a nuclear envelope.
The nucleus is the most prominent feature of a microscope, surrounded by a double membrane called the nuclear envelope. Nuclear pores range in size from 40 to 100 nm, allowing larger molecules to pass between the nucleus and cytoplasm. The nucleolus is a submicroscopic organelle where the ribosomes are assembled. Eukaryotic cells have more than one nucleus, each containing a unique DNA molecule.
The nuclear envelope of eukaryotic cells consists of a double lipid bilayer and is thought to isolate the genetic material inside. In addition to protecting the nucleus, the nuclear envelope is also thought to allow the transport of large molecules, such as proteins. The process of nuclear budding has been implicated in many diseases and increased cellular stress. It also plays a role in protein quality control.
The lining of eukaryotic cells is composed of a ring of proteins called nucleoporins. These proteins stabilize the second positive curvature of the cell wall. They also form the outer ring of the nuclear pore complex. Some of these proteins are also associated with the LINC complex. In addition to forming the nuclear envelope, nucleoporins are also involved in the structure of the cell’s membrane.

















{kind=link}